Research Interests: Non-Natives or Bust!

I am interested in understanding ecological interactions through the lens of non-native species. Species introductions often cause noticeable disruptions to community function and provide "natural" systems on which to test hypotheses about how species respond to novel selective forces. For non-native species, a novel habitat can be a double-edged sword in terms of predators. Typically, when species are translocated to a new environment, they leave behind them a suite of "natural enemies", predators, parasites, competitors, and pathogens. This can be good news for the non-native, an occasionally translates to increased success in the non-native habitat, growing bigger, faster, making more babies. However, the non-native also faces a line-up of unfamiliar threats, including predators to the novel habitat. How do they handle these threats? How do these interactions influence the probability that a non-native species will become invasive?
System the first: Non-native oyster drills scared witless!
 Atlantic Oyster Drills (Urosalpinx inornata)
Atlantic Oyster Drills (Urosalpinx inornata)Credit: E. Grason
During my Master's research, I observed that invasive Atlantic and Japanese oyster drills (Urosalpinx cinerea and Ocenebra inornata) exhibit behavioral plasticity when they detect predation cues from a native crab (Cancer productus) consuming conspecific drills (Grason and Miner 2012). By hiding when the threat is present, drills could avoid being consume by the native predator. This might have contributed to their success as invaders and may also alter post-invasion community dynamics, since drills eat fewer oysters when they are hiding.
So how do they know to hide, and fast? I have observed that alarm cues from injured conspecifics are sufficient to induce the largest response in Atlantic Drills. However, this species does respond to isloated kairomones of an unfed crab, suggesting they do recognize the crab itself. How do they do it? And, more generally, do prey in novel predator-prey interactions prioritize information (e.g., kairomones vs. alarm cues) differently than prey in co-evolved relationships?
So how do they know to hide, and fast? I have observed that alarm cues from injured conspecifics are sufficient to induce the largest response in Atlantic Drills. However, this species does respond to isloated kairomones of an unfed crab, suggesting they do recognize the crab itself. How do they do it? And, more generally, do prey in novel predator-prey interactions prioritize information (e.g., kairomones vs. alarm cues) differently than prey in co-evolved relationships?
System the second: Non-native zombie mud snails festering with parasites!
 Batillaria attramentaria
Batillaria attramentaria
Photo courtesy of Linda Schroeder
and PNWSC
What happens when a non-native species does bring its enemy with it? We often expect that non-natives could experience increased success from the absence of their enemies. But what if they do bring their enemies with them and still succeed? The non-native Asian mud snail Batillaria attramentaria is extremely abundant on the mudflats of Padilla Bay, WA, with abundances recorded as high as 1,500 snails per square meter! Yet the snail is also infected with a co-evolved parasite, for which it is the obligate first intermediate host. This parasite, the trematode Cercaria batillariae, turns snails into zombies by:
Ok, so that's weird. There are billions (really! O'Connor et al. estimated 1.4 billion in a back of the envelope calculation, thats more than there are humans on the earth!) of relatively giant infertile snails wandering around Padilla Bay. But, these snails are about 4 times as dense in Padilla Bay as in their native range. So maybe there is something different about the relationship between snails and parasites in Padilla Bay, that has allowed them to become so abundant. That is, how does the environmental context in which a co-evolved predator prey interaction occurs affect that interaction?
- Castrating their hosts: trematodes consume gonad tissue preventing the snails from reproducing
- Causing gigantism: infected snails resume somatic growth and are as much as 50% longer than uninfected snails
- Inducing migration: apparently, infection cause snails to migrate deeper in the intertidal, presumably to increase the transmission to the next host, crabs or fish.
Ok, so that's weird. There are billions (really! O'Connor et al. estimated 1.4 billion in a back of the envelope calculation, thats more than there are humans on the earth!) of relatively giant infertile snails wandering around Padilla Bay. But, these snails are about 4 times as dense in Padilla Bay as in their native range. So maybe there is something different about the relationship between snails and parasites in Padilla Bay, that has allowed them to become so abundant. That is, how does the environmental context in which a co-evolved predator prey interaction occurs affect that interaction?
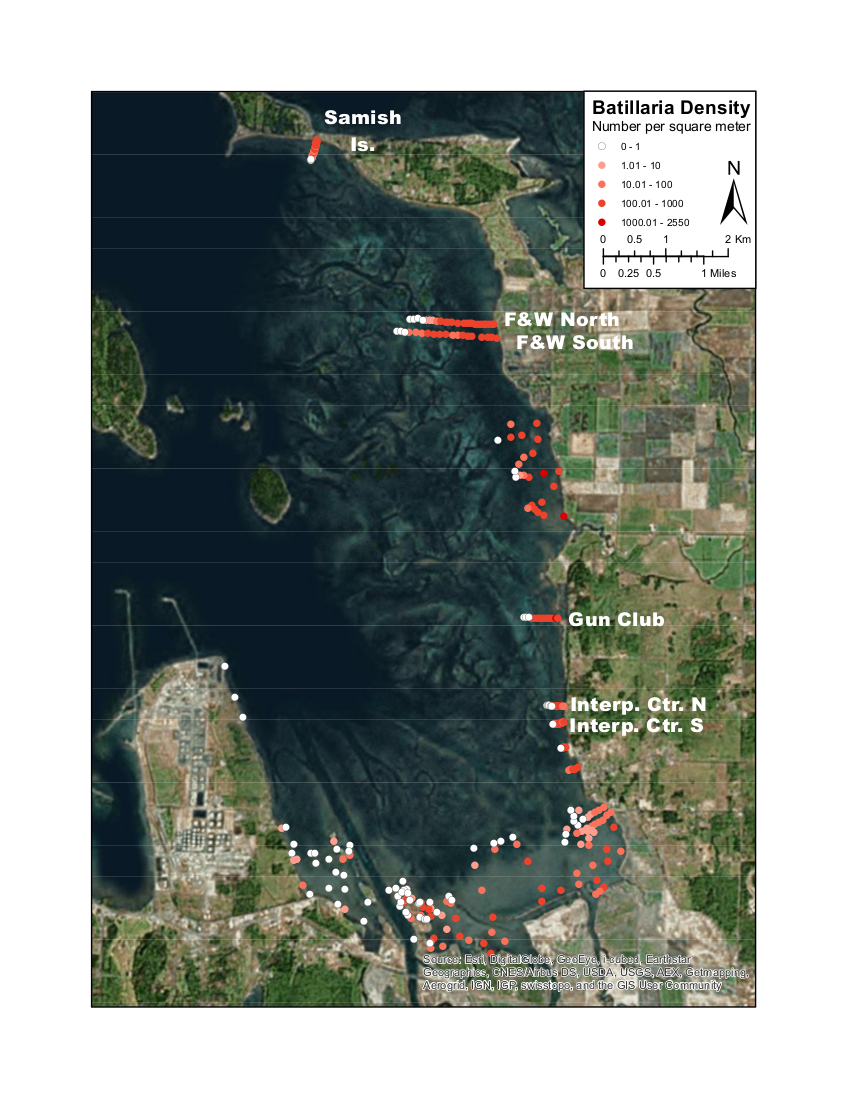
One of the relevant questions was, just where are Batillaria in Padilla Bay, and has the population changed since 12 Billion? I undertook the ponderous metaphysical process of using geospatial modeling to try to update this estimate. Supported by the fantastic support of the folks at the Padilla Bay National Estuarine Research Reserve as part of a three-year NERRS Graduate Research Fellowship, I walked many miles in the mud to generate the density observations in the map to the right. Using a variety of modeling approaches, I estimate that there are between 2.5 and 6 billion snails, though it's really not possible to say weather this evidences a change over time or just a difference in estimation techniques.
Additional work in this system is addressing the following questions:
Additional work in this system is addressing the following questions:
- Does anything actually eat this snail?
- How do Padilla Bay snails compare to snails in a newer invasion, in Willapa Bay?
- Is this snail similar to or different from the oyster drills above in responding to predator cues?
- What is the deal with their parasites?!*